|
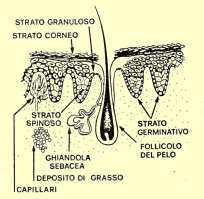
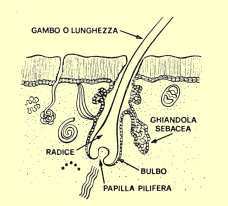
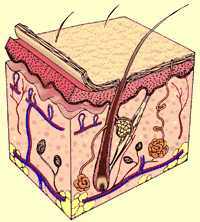
GHIANDOLA
SEBACEA
E' posta lateralmente al follicolo pilifero e ha il compito di produrre il SEBO,
sostanza grassa acida a pH medio 3,5, orientativamente così composta (sulla
superficie cutanea): trigliceridi 30-42%, cere 20-25%, acidi grassi liberi
15-25%, squalene 9-12%, idrocarburi ramificati 4-8%, colesterolo esterificato
2-3%, colesterolo libero 1-1,5%, altre sostanze come di e monogliceridi,
glicerolo ecc 3-5%).
Il sebo ha due funzioni principali:
1) contribuire, con il sudore, alla formazione del film idro-lipidico di
superficie, emulsione acqua in olio che protegge la superficie cutanea dalle
aggressioni chimiche (detergenti, solventi, inchiostri ecc) e batteriche;
2) lubrificare e impermeabilizzare la superficie esterna del capello via via che
questo si allunga.
Il dotto escretore della ghiandola sebacea si apre nella parte superiore del
follicolo in modo da lubrificare il capello prima ancora che quest'ultimo appaia
sulla superficie cutanea. Sul cuoio capelluto la produzione totale di sebo è di
650 - 700 mg nelle 24 ore. Il film idrolipidico, dopo asportazione con
detergente, si ricostituisce in tempi brevi (3 - 6 ore) anche se la ghiandola
sebacea non sembra, al contrario di quanto comunemente affermato, poter
aumentare la propria attività rispetto al normale.
Il sebo non ha la possibilità di spostarsi in modo autonomo lungo il fusto del
capello. Il fatto che i capelli possano nuovamente "ungersi" dopo il
lavaggio deriva, con ogni probabilità, dalla riduzione della tensione
superficiale (determinata da forze fisiche presenti fra le "tegole"
della cuticola di due o più capelli), associata ai continui sfregamenti. La
velocità di migrazione del sebo aumenta se i capelli vengono continuamente
toccati con le mani o pettinati. Inoltre, nei soggetti ai quali si
"ungono" facilmente i capelli, il sebo si presenta più fluido e
quindi con più alta velocità di migrazione.
Nei soggetti con seborrea (vedi) l'abbondanza di sebo provoca numerosi
inconvenienti estetici come adesione delle polveri ambientali, dei residui
cheratinici, delle sostanze di rifiuto veicolate dal sudore e "impiastricciamento"
dei cosmetici applicati.
La produzione delle ghiandole sebacee è controllata dagli ormoni circolanti, in
particolare dagli androgeni di origine gonadica e surrenalica (il più attivo in
tal senso sembra essere l'androstandiolo). Le ghiandole producono attivamente
sebo durante la vita fetale e poi dopo i 9-10 anni. La produzione di sebo
subisce una brusca e notevole riduzione nella donna alla menopausa mentre
nell'uomo diminuisce in genere lentamente dopo i 68 - 70 anni di età.
La
composizione del sebo è la seguente:
1)
acidi grassi liberi 30%
2) gliceridi 29%
3) esteri della cera 20%
4) squalene 11%
5) colesterolo 4%
6) vari 6%
MUSCOLO
PILO-ERETTORE
E' disposto obliquamente a lato della ghiandola sebacea dalla parte dell'angolo
ottuso formato dall'asse del follicolo con la superficie orizzontale
dell'epidermide. Delle due inserzioni, quella profonda è sulla guaina
epiteliale esterna a livello della parte inferiore del terzo medio del follicolo
(sulla zona protuberante) mentre quella superficiale si àncora al derma
papillare per mezzo di fibre dissociate su un'area relativamente vasta .
La sua funzione è, contraendosi, di raddrizzare il pelo
("orripilazione"): l'aria che rimane intrappolata fra i peli si
riscalda a contatto con la superficie corporea ed essendo una cattiva
conduttrice di calore ripara il corpo dal freddo ambientale. Secondo alcuni
Autori la contrazione del muscolo pilo-erettore potrebbe favorire, per
compressione, lo svuotamento della ghiandola sebacea. Il muscolo è assente nei
peli di tipo lanuginoso.
mantenimento del pH normale (5,5) e di una buona idratazione ed elasticità
cutanea.
ANATOMIA
DEL CAPELLO
Nel
capello normalmente sviluppato si distinguono tre parti:
1) una esterna al follicolo, visibile, STELO o FUSTO (spessore medio nell'adulto
65-78 micron);
2) una interna al follicolo stesso, fra l'ostio del follicolo e l'attacco
inferiore del muscolo erettore del pelo, RADICE, "immersa" pertanto
nella cute e quindi normalmente non visibile;
3) una ancora più profonda, alloggiata nella porzione inferiore della parte
profonda del follicolo, BULBO, che contiene, nella parte inferiore (in pratica
il "pavimento") due-tre file di cellule sovrapposte a rapida
riproduzione che costituiscono la MATRICE (originate, come abbiamo visto poco
sopra, dalla guaina epiteliale esterna del follicolo). Queste della matrice sono
le uniche cellule germinative e, pertanto, tagliare corti i capelli non può
"rinforzarli" in quanto quella che si taglia è una parte di fusto,
formata cioè da cellule ormai prive di qualsiasi attività vitale (Il vantaggio
che si può ottenere tagliando corti i capelli è di altro tipo e dovuto alla
minore traumatizzazione con lavaggi, spazzolature ecc); il capello non deve
quindi essere assimilato ad una pianta che necessita o può comunque trarre
beneficio da "potature" periodiche. Le cellule della matrice, via via
che si riproducono, spingono verso l'alto quelle nate in precedenza; durante la
risalita le cellule, in modo analogo a quelle dell'epidermide, elaborano nel
loro interno una proteina, la cheratina, vanno cioè incontro al cosiddetto
"processo di cheratinizzazione" diventando progressivamente sempre più
rigide. Di queste cellule, all'inizio tutte uguali, alcune vanno a costituire la
guaina epiteliale interna (che tende a cheratinizzarsi e quindi ad indurirsi in
anticipo rispetto al pelo in modo da "modellarlo"), altre le tre parti
strutturali del capello vero e proprio (cuticola, corteccia e midollo). Subito
al di sopra delle cellule delle matrice sono presenti alcuni melanociti che
hanno il compito di "colorare", immettendovi la melanina, quelle
cellule che diventeranno la "corteccia" del capello.
Secondo G. Cotsarelis e collaboratori (University of Pennsylvania e New York
University School of Medicine) le cellule germinative risiederebbero non nella
matrice ma nella già precedentemente ricordata "zona protuberante";
l'Autore riferisce che quando si arriva alla fase catagen la matrice vera e
propria degenera e la papilla rimane connessa al bulbo solo una lunga colonna di
cellule epiteliali; in seguito la papilla risale attivando le cellule
germinative della zona protuberante che migrano verso il basso colonizzando
nuovamente la matrice e dando inizio al nuovo ciclo del capello; questa presunta
"nuova sede" delle cellule germinative potrebbe spiegare i parziali
insuccessi finora ottenuti relativamente alla possibilità di ottenere, in
laboratorio, delle colture cellulari capaci di dare origine al pelo: sarebbero
state infatti utilizzate le cellule sbagliate!
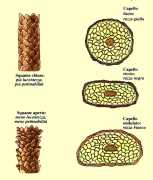
Secondo Rook capelli e peli ricci avrebbero origine da follicoli pilosebacei
curvi con asincronia fra tasso di crescita e di cheratinizzazione delle due
pareti opposte della guaina epiteliale interna.
STRUTTURA DEL CAPELLO
3)
MIDOLLO, interno, formato da cellule arrotondate, disposte a colonne,
generalmente separate fra loro da spazi d'aria (l'aria trattenuta ha funzione
determinante per la protezione dal freddo e per questo motivo il midollo supera
negli animali il 50% dello spessore totale del pelo mentre è scarsamente
rappresentato e talora assente nella specie umana).
COMPOSIZIONE
CHIMICA DEL CAPELLO
All'analisi chimica del capello i costituenti principali, oltre all'acqua, sono:
cheratina, lipidi, minerali e pigmenti.
Cheratina:
è una proteina contenuta principalmente nella corteccia (della quale ne
costituisce la massima parte); gli aminoacidi presenti nella cheratina sono 18
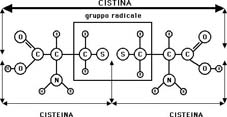
ma, in maggiore quantità, ritroviamo la cisteina, la cistina (derivata dalla
condensazione di due molecole della prima con perdita di acqua e formazione di
legami disolfuro), la serina, l'acido glutammico, la glicina, la treonina, l'arginina,
la valina, la leucina e l'isoleucina.
La
cheratina presente in maggiore quantità nel capello è quella alfa, fibrosa, a
basso contenuto di zolfo, con Peso Molecolare di circa 45.000, insolubile in
acqua. Le catene polipeptidiche sono rese stabili da tre tipi di
"ponti": legami idrogenati (conferiscono solidità), ponti fra catene
acide e catene basiche (si rompono con gli acidi forti) e ponti disolfurici
(quando sono lesi, ad esempio nelle "permanenti", il capello si
arriccia). La cheratina può essere deformata con il vapore acqueo ("messa
in piega"). Il processo di cheratinizzazione, in particolare quello dei
peli, è regolato da ormoni, vitamine, fattori genetici e metabolici e sembra
legato al metabolismo del colesterolo e alla sua esterificazione con acidi
grassi sintetizzati dall'epidermide. Ne consegue che carenze dietetiche e/o
difetti enzimatici delle vie di sintesi del colesterolo e degli acidi grassi
possono provocare una cheratinizzazione anomala con conseguenti difetti
strutturali del fusto.
Lipidi:
costituiti da trigliceridi, cere, fosfolipidi, colesterolo, squalene ed acidi
grassi liberi sono quantitativamente documentabili con estrema difficoltà in
quanto in massima parte derivati da quelli del sebo.
Minerali
(oligoelementi): rappresentano una componente essenziale dei sistemi
proteico-enzimatici. Esiste una correlazione diretta fra la quantità di
oligoelementi presenti nel sangue e quelli presenti nel capello.
a) ferro (media 4-12mg/gr): è più abbondante nei capelli rossi rispetto a
quelli biondi e neri.
b) magnesio (media 30-45mg/gr): è più abbondante nei capelli di colore nero
(fino a 170mg/gr). Catalizza la trasformazione dell'ATP in cAMP, in seguito alla
attivazione della adenilciclasi, e la degradazione dell'cAMP in 5-AMP ad opera
della fosfodiesterasi (vedi).
c) zinco (media 150-180 mg/gr): è indispensabile per la corretta attività
delle cellule germinative della matrice. In sua carenza il capello si
indebolisce e rallenta il ritmo di crescita.
L'eccesso di zinco determina carenza di rame a causa dell'induzione dell'enzima
tioneurina che, a livello intestinale, lega il rame in modo preferenziale
rallentandone o impedendone l'assorbimento.
d) rame (media 16-50mg/gr): è indispensabile per catalizzare la conversione
della tirosina a DOPA (nel processo di sintesi della melanina) e per consentire
l'ossidazione della cisteina in cistina con formazione dei ponti disolfuro.
e) piombo (media 10-30mg/gr): è più abbondante nei capelli di colore castano.
Dato che la quasi totalità del piombo presente nell'organismo deriva da quello
esogeno e poiché il capello ne rappresenta la sede principale di accumulo, la
sua misurazione a livello della parte distale del fusto viene utilizzata per
valutare la presenza di questo metallo nell'inquinamento ambientale.
La
carenza di proteine e/o sali minerali sarà evidenziata al microscopio da un
fusto sottile associato a bulbi molto piccoli (se il capello è
costituzionalmente sottile i bulbi appariranno invece normali).
Pigmenti
: sono rappresentati dalle melanine (sostanze colorate), presenti nel pelo in
forma diffusa o granulare. Sono insolubili in acqua, solubili negli acidi forti,
decolorabili con acqua ossigenata.
I melanociti, utilizzando la tirosina (che è un aminoacido, cioè un
costituente delle proteine) come precursore, sintetizzano due principali tipi di
melanina: l'eumelanina, scura e presente nei capelli neri e la feomelanina, più
chiara e presente nei capelli dorati, biondi o rossi .
I
principale elementi chimici presenti nel capello sono:
1)
carbonio 45%
2) ossigeno 28%
3) azoto 15%
4) idrogeno 6,5%
5) zolfo 5,2%
Sono
presenti inoltre diversi oligoelementi fra cui si possono determinare con metodi
complessi (Mineralogramma) la presenza di: Ca, Mg, Sr, B, Al, Si, Na, K, Zn, Cu,
Mn, Fe, Ag, Au, Hg, As,Pb, Sd, Ti, W, Mo, I, P, Se
Le
percentuali degli oligoelementi presenti nei capelli sono una caratteristica
soggettiva di ciascun individuo come le impronte digitali.
L'80%
circa del peso del capello è dovuto alla presenza di proteine (polimeri di
amminoacidi) fra cui la principale è la cheratina composta da 18 amminoacidi.
I
principali amminoacidi che compongono la cheratina sono:
1)
cistina 17,5%
2) serina 11,7%
3) acido glutamico 11,1%
4) treonina 6,9%
5) glicina 6,5%
6) arginina 5,6%
|

|























